The following article was first published in Wild Earth, Volume 6, #1, 1996, pp 37-43. Certain corrections, usually indicated by brackets, have been introduced into the text. Also, the original published version erroneously contained a large duplicated section, which is eliminated here.
Biodiversity: Central Appalachian Plant Distributions and Forest Types
or What a Walk in the Woods Can Tell You
by Robert F. Mueller
ABSTRACT
Occurrences of plant species and forest types in the Central Appalachians are related to elevation, soil acidity and moisture content. Simple observations and survey methods are featured. Time rate of change (kinetic) and equilibrium criteria for forest succession are discussed.
PATTERNS IN THE WOODS
If knowledge is power, ignorance is fatal when it comes to our threatened native ecosystems and their beleaguered defenders. The enthusiastic nature lover may be awestruck and inspired by nature's grandeur, but in the absence of real knowledge and detail, the experience may be little more than a green emotional blur or an inchoate impression of sights and sounds not likely to arm one effectively.
We don't need to be card-carrying professionals to know the forest. Professional foresters are demonstrating every day that their training may actually be counter-productive. Rather, we can begin in our own way with identification of common trees and animals, and progress gradually to less eye-filling and more difficult species. Virginians for Wilderness have for some years been humbly packing their field guides and manuals into the woods, trying to get on familiar terms with the bewildering Appalachian diversity. We are using this hard-won familiarity to understand the processes at work in forming both natural and technologically influenced biologic communities. To this end, we've made traverses up as many ranges, peaks and "knobs" as possible, collecting data on the way up and down. Such climbing traverses have the advantage of taking us over most of the representative terrain types, ranging from low elevation coves and major riparian zones through mid-slope topographic convexities and concavities to the most exposed summits. We also try to extend our studies to as many as possible of the unusual biologic communities such as isolated mountain bogs, glades and barrens which are frequently home to rare species and highly disjunct populations. These unusual habitats are important because they may contain species at the limits of their ranges, so that the conditions rendering their occurrence possible are most starkly displayed. We have noted that many northern species (e.g. Balsam Fir and Buckbean), which are here at their southernmost stations or nearly so and are of very restricted occurrence, nevertheless flourish and reproduce prolifically at these stations as long as conditions remain favorable and there are no human-induced disturbances. At issue here is the concept of equilibrium as distinguished from change with time or plant succession.
The effect of elevation on species ranges and forest types was discussed extensively by Braun (1950). For example, she mentions the elevational limits of Tuliptree and other species and notes the occurrence of index species such as Maianthemum canadense, Clintonia borealis and Trillium undulatum for northern / high elevation habitat conditions. Here an attempt is made to extend this approach by graphical approximations.
Figures 1, 2 and 3 summarize the results of field studies and literature surveys on certain habitat indicator species, forest type distributions and environment-sensitive plant communities in the vicinity of 38o N latitude. In these figures, elevation above sea level is straightforward since it is easily determined. However, pH is much less well defined not only because of instrumental problems, but because there is seldom a single pH reading for a column of soil at any given location. In many cases, pH may vary by almost 3 units in a 50 centimeter column of soil (Armson 1977). In the eastern US, soils generally show lower pH (more acidic) values in leached surface horizons because precipitation is acid even in the absence of anthropogenic pollutants. This is true even in soils developed on limestone bedrock: horizons may be quite acidic near the surface but near neutral (pH=7) or alkaline in contact with limestone at depth. Some tree species (e.g. Black and Red Spruces and Arborvitae), though, have very shallow root systems, particularly in bogs, so the surface pH may for such trees be a reliable indicator of the effective pH under which these trees developed. Also, a number of species (e.g. Chinquapin Oak, various ferns, mosses and lichens) frequently are in direct contact with rock surfaces that contain pH-determining minerals such as calcium carbonate. Although not precise, then, pH values can be used to help explain floral and faunal occurrences. Soil moisture, too, is a difficult parameter, but relative magnitudes may be estimated by comparison with the end points of standing water and the observed driest sites.
Figure 1 shows the most prevalent ranges in elevation and pH in the Central Appalachians with plotted occurrences of some common trees, shrubs and herbaceous species. Some major forest trees-including Tuliptree, Black Gum, Chestnut Oak and Sassafras-appear to be relatively insensitive to pH, but are limited by elevation, hence their limits are indicted by horizontal lines. Thus Auten (1945) found no correlation of site quality with calcium, magnesium, phosphorous, potassium or soil reaction pH for Tuliptree. Such species are commonly observed growing over a wide range of conditions, though Tuliptree and some others are very moisture dependent. Red Spruce (Picea rubens), the common high elevation conifer on the Allegheny Plateau, appears to be limited by pH and generally grows in podzolic soils with pH from 4 to 5.5 (Fowells 1965). Although the lower limit of this tree is shown as a horizontal line, this limit may vary in elevation with pH and certainly does with moisture conditions. The same applies to species such as Mountain Holly (Ilex montana) and Canada yew (Taxus canadensis), which have less well known pH ranges but are placed in the figure based on field observation of general habitat. Quite generally, in this figure upper and lower elevation limits of species are indicated by downward and upward pointing arrows respectively.
Some species, such as Ilex montana, represent ambiguous cases. Although this species is characteristically encountered at high elevations in the Central Appalachians, it forms part of a complex that extends into the Piedmont and Coastal Plain and has been referred to as Ilex ambigua (Radford et al 1964).
In this scheme, Partridge Berry (Mitchella repens) is a special case. This little evergreen creeper on the forest floor, with the shiny leaves and red berries, is a conspicuous member of acid soil communities from sea level to the highest Allegheny spruce forests (indicated by the vertical line). However, its precise and full pH range is uncertain (as shown by the horizontal arrow with a question mark).
Occurring in much the same environment as Partridge Berry, but usually confined to high elevations, are the four northern herbaceous species, Canada Mayflower (Maianthemum canadense ), White Wood Sorrel (Oxalis montana ), Yellow Clintonia (Clintonia borealis) and Bunchberry (Cornus canadensis ). In figure 1 these species are shown with upward pointing arrows indicating the approximate lowest common elevation of occurrence. Cornus canadensis is very rare in the Central Appalachians. Maianthemum canadense is, in contrast, quite common at intermediate to high elevations, while Oxalis montana and Clintonia borealis appear to be of intermediate frequencies at higher elevations. These four species are written slanting upward to the right to reflect an inferred interaction with pH. By Le Chatelier's [ Principle ], a stressed system will react in such a way as to relieve the stress. Thus an acid-loving northern or high elevation plant might be expected to relieve the stress of warmer temperatures at low elevations by seeking a more acid environment. Although this [principle] is usually applied in chemistry, plants are of course complex chemical systems, and if other factors remain constant, it should enter here also.1
A similar approach is applicable to the assemblage of plants at the lower right in Figure 1. These plants range from the common Maidenhair Fern (Adiantum pedatum) to the Chinquapin Oak (Quercus muehlenbergii) in lime-rich environments at pH values near neutral. Lance-leaved Buckthorn (Rhamnus lanceolata), Walking Fern (Camptosorus rhizophyllus ) and Chinquapin Oak characteristically occur very near or on limestone or lime-rich outcrops, and the latter tree is seldom found in environments with pH lower than 7 (Reynolds and Potzger 1953). The species written slanting up to the right are all southern or temperate species so that the stress of cooler (high elevation) environments would be expected to be relieved by higher soil pH values.
Arborvitae (Thuja occidentalis) is a special case because it is a distinctly boreal species that in the north is found in swamps, some of which are quite acid. In fact its leaf litter has a pH ranging from 4 to 4.9 (Fowells 1965). Furthermore, Arborvitae seedlings usually develop in rotten wood, decayed litter, peat and moss, which are characteristically acidic substrates. In the north and intermediate latitudes, Arborvitae also occurs in alkaline environments such as limestone outcrops and calcareous wetlands. However, in the Central Appalachians it is with rare exception found on or near limestone or carbonate-bearing rocks, usually at moderate elevations. It appears that the Central Appalachians contain few high elevation bogs suitable for Arborvitae. Apparently, then, the stability field of Arborvitae is constrained to alkaline environments at low latitude low elevation locations but is expanded to include both alkaline and more acidic environments at cool high latitude as well as hypothetical high elevation locations.
It must be remembered that the species shown in Figure 1 are only a small fraction of those that respond to differences in elevation and pH. Conversely, many species of very wide occurrence show little obvious response to these parameters. Red Maple is a striking example which, though it shows some aversion to high pH values, is present in habitats ranging from southern swamps to the fringes of the boreal forest, including all elevations in the Central Appalachians. Other common trees of wide distribution in many habitats are White Oak, White Ash and Black Cherry, although these are generally excluded from swamps and areas of low soil fertility.
Well recognized forest types of the Central Appalachians as related to geographic distribution and elevation are shown in Figure 2. Nomenclature generally follows that of Braun (1950) but has been modified to distinguish well segregated Red Spruce Montane Forest which she included in her "Hemlock-White Pine-Northern Hardwoods" type. Major segregates of the Oak-Chestnut type are also shown. As indicated previously (Mueller 1994), these variations in forest type appear to reflect, not only temperature and rainfall differences, but also a systematic variation in degree of cloudiness from east to west. In Figure 2 the full zig-zag lines indicate inter-fingering or blending of different forest types in both horizontal and vertical ranges. The horizontal dashed line that divides the Oak-Chestnut region distinguishes the dominantly Northern Red Oak "orchard" subtype of high elevations from the more extensive forests of lower elevations dominated by combinations of Chestnut, White, Scarlet, Red and Black Oaks. The dividing line between the two subtypes is the upper limit of Chestnut Oak as shown in Figure 1. Similarly, the triangular area indicates the cove type forests within the regional Oak-Chestnut type. These cove forests are similar to the mixed mesophyte forests farther west, but seldom contain Yellow Buckeye and certain other species present in the latter. In the highest versions of these cove forests, usually above 2500 feet in elevation, Tuliptree and some other southern species drop out. While both the Valley and Ridge and Blue Ridge Provinces are characterized by Oak-Chestnut upland forests and similar cove forest vegetation, there are differences in detail (Mueller 1994). Neither Figure 1 nor 2 take account of aspect or differential exposure to sunlight as related to slope. This is generally difficult to detect in the Central Appalachians given all the contributing factors, and needs more observation.
To illustrate the response of vegetation to soil moisture and pH, it helps to include some specialized communities, Figure 3 shows the most water saturated plant community, the peat bog. These bogs, as typified by those on the Allegheny Plateau, are dominated by a variety of mosses, particularly Sphagnum, and lichens such as Cladonia (so-called "Reindeer Moss") and may contain acid-loving flowering plants such as sundews and cranberries. Associated trees are usually spruce, Balsam Fir and Eastern Larch. Quite different is the flora of calcareous fens and marshes, which are far richer in available nutrients, and have a greater diversity of flowering plants-including some, such as Swamp Lousewort (Pedicularis lanceolata), Prairie Loosestrife (Lysimachia quadriflora) and Large-leaved Grass of Parnassus (Parnassia grandiflora ), that are virtually confined to a calcareous environment and are thus highly diagnostic of it. While conditions on the dry end of Figure 3 are not as well defined as on the wet end, communities that occupy it are quite distinctive and certain combinations may immediately be excluded by reference to this figure. For example, one would rarely, if ever, expect to see a peat bog surrounded by Chinquapin Oak or in contact with a calcareous fen.
USING THE KNOWLEDGE
I have tried to illustrate how it is possible to find order and consistency in nature by making use, primarily, of walks in the woods. It is, of course, desirable to supplement this information whenever possible by more detailed studies such as biologic transects and lab work. Frequently, however, the time frame of the forest activist doesn't permit exhaustive studies, for which facilities in any case may be lacking. There is an urgency to our work, forced by a multitude of threats, that makes it necessary to get things done "now."
Noss (1992) stated that "No substitute exists for detailed on the ground knowledge of the ecology and natural history of the region." This study, based as it is on thousands of observations of plants in relation to their environments, represents an effort to gain such knowledge. Let's now consider a few ways in which such knowledge can advance attempts to protect forests. Since many of the plants discussed are indicator species, they provide evidence of such features as karst topography, including caves and mineral "licks," which may otherwise be concealed. We ought not let the mining industry, which regularly uses plants as prospecting tools, get ahead of us on this. Karst topography harbors some of the rarest species known. The same reasoning applies to acidic or other unusual or extreme environmental parameters. Reserves designed without this type of information might miss critical or even keystone species and communities and so preclude long-term viability. Information of the type presented here is also necessary to evaluate frequently misleading data presented by the U S Forest Service and other land management agencies. Often these agencies make use of highly artificial forest type designations, which may list species incompatible with the environment of stands thus designated. In the Monongahela National Forest, for example, "Black Cherry-White Ash-Yellow Poplar (Tuliptree) " forest type is occasionally attributed to areas in which Tuliptree is excluded by elevation. Similarly, some high elevation forests in the George Washington N F have been erroneously classified as Scarlet Oak type. Other applications of field classification of selected species or communities with the physical environment will no doubt occur to discerning readers as they explore areas they have adopted for protection.
Although the results of this study are strictly applicable only to the vicinity of 38o N latitude, they may be extrapolated using the approximation that each 1000 feet change in elevation corresponds to a 300 mile change in latitude. Thus the upper limit of a tree such as Chestnut Oak would be raised a little more than 300 feet a hundred miles south of latitude 38o N. However, such extrapolation should also take into account east-west climatic [ element ] variation such as cloud cover, changes in aspect due to changes in ridge orientation, and other factors that may enhance or counter lapse rate.
. A result that may follow from floral-and faunal-distributions such as those illustrated in Figures 1-3 is an improved picture of dynamic equilibrium in these forests. The concept of dynamic equilibrium may be defined as the tendency of a perturbed system to return to a pre-existing state. In natural systems such perturbations are generally natural disturbances. In the mineral systems of rocks and soils, equilibrium is sometimes difficult to prove and may be present to varying degrees and confined to restricted volumes (Mueller and Saxena 1977 ), pointing to the importance of scale. In chemical systems of all kinds it is common to find states of metastable equilibrium in which a system is in a relatively stable, but not the most stable state. In addition, most natural systems are "open" and equilibrium is superimposed on a steady state in which energy and materials exchange with the surroundings. In such systems equilibrium can prevail only if the rate of the equilibrating process is greater than that of the exchange. Quite generally, deviation from equilibrium is the province of the science of kinetics, or the time rate of change of systems.
Braun (1950) discussed some criteria for forest equilibrium (climax) : (1) accordance of canopy and understory, (2) tendency of the same climax to develop as a result of unlike succession, (3) occupation of topographically mature sites, (4) equilibrium between soil and occupying vegetation and, (5) climax similarity over a large area. Biologists now reject point 3 as a criterion, since they've observe that a climax may be developed on topography as new as glacial moraines. Also, point 1 needs qualification. Reproduction in areas in which the canopy has been opened temporarily may be quite different from the overstory. Also, in some cases [ accordance ] is not necessary (Chestnut Oak seedlings under Chestnut Oak canopy), in other cases it is not sufficient ( aspen sprouts under aspen), to establish equilibrium. Nonetheless, in the case of tolerant seedlings and saplings under mature trees of the same species, evidence may be compelling. In general, Braun's ideas are in agreement with the definition of dynamic equilibrium.
To the above criteria must be added the recovery of climax forests from large scale disturbances such as disease and, in particular, the historic episode of logging and agricultural abuse. One of the best examples is the ongoing return of high elevation Red Spruce in the Allegheny Plateau's northern hardwoods which replaced them after logging and subsequent fires. Of course, where soils have been badly degraded, changes in forest type occur on a more permanent basis (Braun 1950). In some places, as at West Virginia's Cranberry Glades, the establishment of the regional climax is well documented by the pollen record (Core 1955).
The largest disturbances to which the Appalachian forests were subjected are the ice-age and post-ice-age migrations. The data of Delcourt and Delcourt (1993) suggest that biotic responses approach dynamic equilibrium in the Central Appalachians beginning about 8000 years before present for cool temperate deciduous forests, but that the mixed conifer-northern hardwoods north of 44o N latitude have not attained this state even today. Naturally, this conclusion addresses a more or less crude equilibrium that ignores minor forest migrations associated with temperature fluctuations, particularly around 7000 years ago. If this conclusion of equilibrium on a broad geographic scale is adopted, it follows that elevational zoning as indicated by Figures 1 and 2 is also evidence of equilibrium since it recapitulates, with modifications, the latitudinal zonation. Again, where disturbances (primarily anthropogenic) have intruded, as in the Allegheny spruce forests, the tendency is for the forest to return to the equilibrium type. These conclusions are in general agreement with Braun's criteria 2, 4 and 5, as well as the general concept of equilibrium.
Although data on forest distribution and zonation suggest equilibrium, "proof" of it may be out of the question, since disturbances both natural and human-induced are so prevalent and the response/recovery times often so long as to preclude all but crude correspondence. This is illustrated by certain secular deviations from prevalent climaxes in the Central Appalachians. For example, in some places Red Spruce forms an understory under old-growth or mature oak-rich upland forest, as in the Fanny Bennett Hemlock Reserve in the Monongahela National Forest, where it cannot be easily attributed to recovery from the human-induced episode of deforestation and fires. Here, though, the change may result from recent fire control measures and so represent a shift of equilibrium under a modified disturbance regime. An alternative explanation is that such expansion of the spruce forest represents a recent change to cooler and moister conditions.
To reiterate, the results of this study are tentative and approximate. The limits on species distributions by elevation, pH and soil moisture are not absolute. Fairly common types of variation are where microhabitats or microclimate imposed by the geology or topography make it possible for high elevation/northern species to flourish at low elevations or low elevation species to extend to higher elevations. An extreme case is Ice Mountain in Hampshire County, West Virginia, where a boreal plant community exists at only 700 feet above sea level as a result of persistent ice accumulation in a talus slope. Quite generally, disjunct species are more frequent in seeps, bogs or other wetlands, where water seems to have a buffering effect (Mueller 1994). The reader is encouraged to refine and expand these data, not only in the Central Appalachians, but in other areas where the same species occur. Only through such painstaking efforts can we create the scientific basis for the reserve system we need so urgently (Noss 1992).
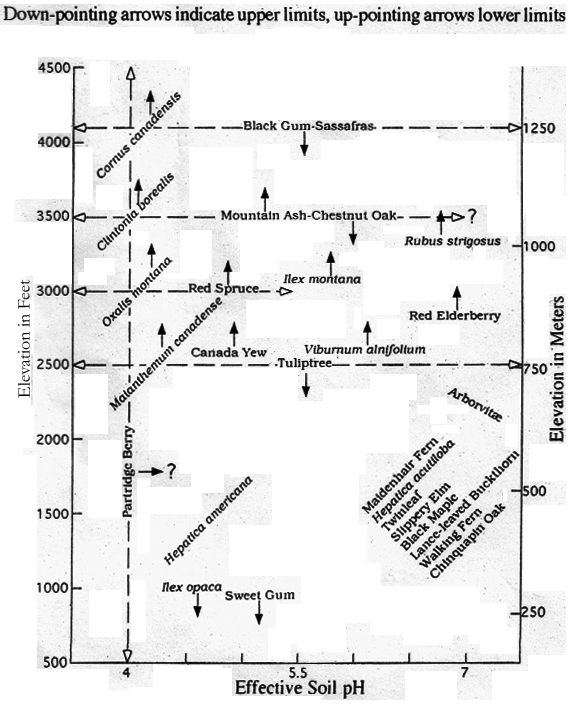
Figure 1: Central Appalachian Habitat Indicator Species as a Function of Elevation and pH at 38o N Latitude
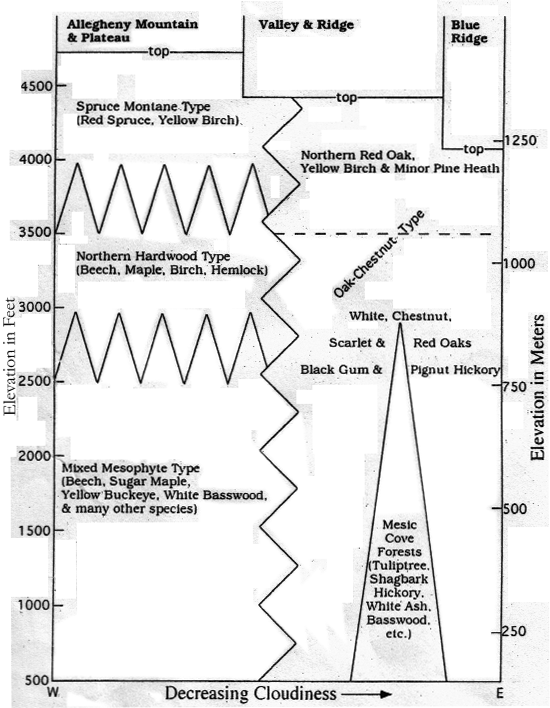
Figure 2: Major Central Appalachian Forest Types at 38o N Latitude
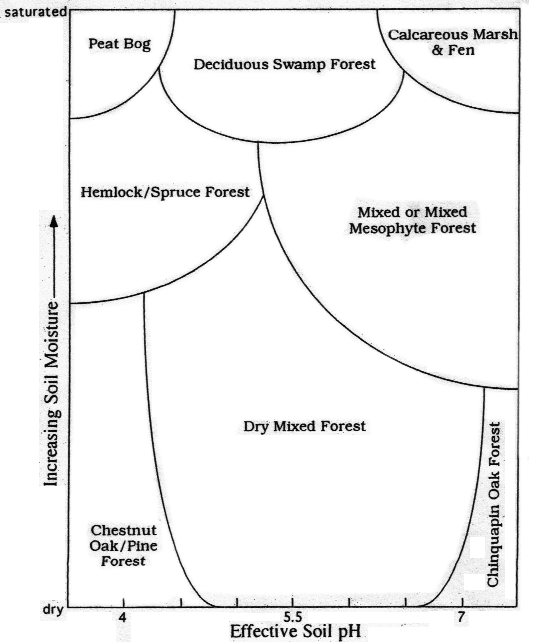
Figure 3: Central Appalachian Plant Communities as a Function of Soil Moisture and pH at 38o N Latitude
Acknowledgements
The writer appreciates the assistance in the field as well as the continuous intellectual stimulation of Mike Jones, Steve Krichbaum and Gus Mueller. Gus Mueller's assistance with the figures was also invaluable. Particular gratitude is due Dr. Robert Hunsucker, who has advanced our efforts greatly through his knowledge of the Appalachian flora and fauna. The partial funding of this work by Human-i-tees is gratefully acknowledged.
References
Armson, K. A. (1979) Forest Soils, Properties and Processes. University of Toronto Press, Toronto, Ontario
Auten, John T. (1945) "Prediction of Site Index For Yellow-Poplar From Soil and Topography." Journal of Forestry 43, pp 662-668.
Braun, E. Lucy (1950) Deciduous Forests of Eastern North America. Macmillan, New York.
Core, Earl L. (1955) "Cranberry Glades Natural Area." Contribution No. 74 from the Herbarium of West Virginia University. Wild Flower 31, pp 65-81.
Delcourt, Paul A. and Hazel L. Delcourt (1993) "Paleoclimates, Paleovegetation and Paleofloras during the late Quaternary." In Flora of North America North of Mexico. Flora of North America editorial committee, Vol. 1, Chapter 4, pp 71-94. Oxford University Press, New York.
Fowells, H. A. (1965) Silvics of Forest Trees of the United States, Agricultural Handbook 271. U. S. Dept. of Agriculture, Washington, D. C.
Langridge, J. (1963) "Biochemical Aspects of Temperature Response in Ants." Annual Review of Plant Physiology, pp 441-426.
Mueller, R. F. (1994) "Central Appalachian Forests, A Guide for Activists." Wild Earth 4, #3, pp 37-49.
Mueller, R. F. and S. K. Saxena (1977) Chemical Petrology with Applications to the Terrestrial Planets and Meteorites. Springer-Verlag, New York.
Noss, Reed (1992) "The Wildlands Project Land Conservation Strategy." Wild Earth, Special Issue, pp 10-25.
Ogle, Douglas W. (1989) "Barnes Chapel Swamp: An Unusual Arbor-vitae (Thuja occidentalis) Site in Washington County, Virginia." Castanea 54 (3) pp 200-202.
Radford, Albert E., Harry E. Ahles and C, Ritchie Bell (1964) Manual of Vascular Flora of the Carolinas. University of North Carolina Press, Chapel Hill, N.C.
Reynolds, William and J. E. Potzger (1953) "Distribution of Quercus muehlenbergii in Indiana." Butler Univ. Bot. Studies 10, pp 71-79.
Robert F. Mueller [727 Stingy Hollow Road, Staunton, VA. 24401] is a retired NASA scientist, naturalist, and leader of Virginians for Wilderness.