A perspective of terrestrial ecosystems is developed in which plants are regarded as occupying stability fields in volumes of multidimensional space formed by physical variables such as temperature and chemical composition of the substrate in addition to the familiar space axes. Examples of observed stability fields of some forest trees are given as are others involving herbaceous species. Inclusion of climatic, topographic and biologic factors are discussed, and the importance of microhabitats is stressed. Applications to forest ecology and floral diversity and type in old growth forests is made based on new data. This perspective has the advantage of providing a unified approach to a variety of data. Because it incorporates the methods and subject matter of mineral and physical chemistry it promises to broaden the scientific basis of conservation biology and fortifies ecosystem protection as well.The Model
How we see the forest, our mental perception, is important, not only in recognizing threats to it, but also in providing tools to protect it. The world in which we live has a dimensionality that we move through in all we do. To most of us this dimensionality is revealed in our spacial coordinates of latitude, longitude and elevation or equivalents. However, our perspective increases greatly when we make the generalization from three dimensional space to include axes such as time, temperature and a multitude of others that an in-depth study of nature demands. During my years of research in mineral chemistry I was essentially exploring the multidimensional space of Earth's crust along temperature, pressure and chemical composition axes.[1] So it was inevitable that I have come to recognize a similar, in some cases even the same, dimensionality in ecosystems. However, there is a difference too. While the determinants in mineral chemistry are thermodynamic, a stable forest is not in thermodynamic equilibrium and ecologic systems are so complex that they force us to fall back on more general principles of dynamic change and stability. However I firmly believe that the approach that has proved indispensable in the study of mineral systems provides a template to treat ecosystems as well. Furthermore, life processes clearly merge with those of physical chemistry in these systems.General Forest Ecology and Old Growth ApplicationsA useful adaptation from mineral systems to forest ecosystems is the stability field concept, which comes ultimately from physical chemistry. Quite generally a system is stable when it returns to a pre-existing state after a perturbation. However there is also a more restricted form of stability, namely metastability, in which a system is in a relatively stable but not the most stable state. In mineral systems this corresponds to a low but not the lowest Gibbs free energy state. Examples are supercooled raindrops and the high magnesium shells secreted by certain marine animals. A non-thermodynamic example (which may however embody thermodynamic elements) from silviculture is a Red Spruce tree planted below its elevational or latitudinal limit or a Black Walnut planted above. In either case the tree may thrive but will not reproduce. Plants may also find themselves in metastable situations as environmental conditions change.
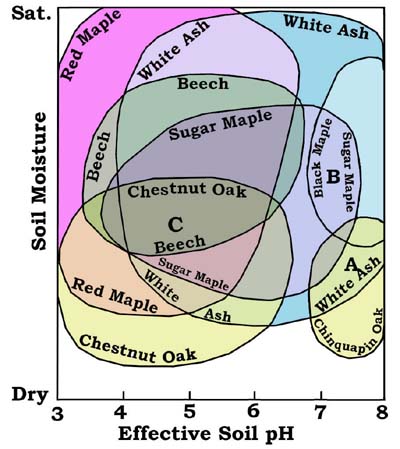
Fig. 1-Tentative overlapping tree stability fields with resulting associations (A, B, C) as a function of field position. Concept by RF Mueller, graphic by the Gus.A stability field is a volume of multidimensional space in which a species is stable or in some cases metastable. Soil moisture and pH (acidity) are two of the most useful axes in plotting the stability fields of many forest species. This is a direct consequence of the almost universal importance of humidity and acidity in biochemical processes (Edsall and Wyman, 1958). Figure 1 illustrates the stability fields of some common and less common trees in terms of these axes. Although the soil moisture axis shows only relative field boundaries along it, these are defined by recognizable dry and saturated limits. The figure shows how associations arise from overlapping fields and one reason why some species are less common than others. Although the figure is based on extensive field data, it is still tentative and subject to correction. Because the figure is a two dimensional slice through multidimensional space, it is subject to variations along other axes, such as for example in the Calcium/Magnesium ratio. It must be kept in mind that such figures express necessary but not sufficient conditions for the presence of a given species, which may depend on other factors such as available seed sources or succession in which different species occupy the same or slightly different stability fields at different times. Another caveat is the possible alteration of stability relations by the species themselves; which happens when Hemlock acidifies the upper soil layers by needle drop. In addition genetic variation may expand a species' stability field. Despite such complications --which I have not exhausted above-- certain associations are clearly excluded. The use of compatible and incompatible species -- also a direct carryover from mineral chemistry -- provides a readily available and practical means of rapid assessment and characterization of forest tracts. As illustrated in the figure, which is only one of many possible, certain tree species do not normally occur together since their stability fields do not overlap, and if such incompatible species are found in close proximity, a steep gradient along some axis is indicated, or alternatively, other axes may be involved.
After this paper was written Reed Noss drew my attention to a book by G. E. Hutchinson ( 1965 ). In this interesting work the author proposed a model in which a volume of multidimensional space ( "hyperspace" ) represents a "fundamental niche" of a species. Although he did not explicitly treat forest ecosystems and virtually confined his examples to animals, his model is clearly congruent with that proposed here. Of particular interest is his treatment of competition between similar species and which may be extended to forest plants as well. Despite the importance of this concept of a niche, the alternative of stability field is retained here since the latter is not limited to organisms but also applies to the mineral substrates upon which organisms depend and thus unifies more diverse phenomena.
Moisture, acidity, temperature and other axes also reveal themselves in sequences of herbaceous and woody plants with different stabilities along these axes. In some cases "dropout sequences" occur in which a number of species occur together at some temperature or pH, for example, but drop out in succession as these parameters either increase or decrease. Thus Teaberry (Gaultheria procumbens), Rattlesnake Plantain (Goodyera pubescens), Rattlesnake Weed (Hieracium venosum) and Spotted Wintergreen (Chimaphila maculata) may occur together at high acidity (pH=3 to 4) ranges but drop out in the approximate order they are written in above as acidity decreases. Preliminary observations suggest a number of such sequences and one involving temperature was previously presented (Mueller, 1996). An analogous system was introduced by Küchler (1972) in his analysis of "phytocenoses" or natural plant communities. In this system overlapping species ranges were used to fix such parameters as pH for the community.
Despite differences, there is a close link between the inorganic mineral world and ecosystems, a link far closer than recognized in much current research. Thus many articles on natural biologic systems and forests in particular fail to specify the nature of the mineral substrate. While most rocks and their contained minerals were formed or modified deep within Earth's crust, their exposure on the surfaces drives them toward an equilibrium suitable to those conditions. And it is this trend toward equilibrium -- in this case thermodynamic -- that creates, with the participation of climate and organisms, the substrate for terrestrial organisms and by extension aquatic ones as well. Thus it is hardly surprising that plants have evolved specific relations with various substrates and that these relations extend to the bedrock and derived materials. Carbonates (limestone, dolomite, and some other sedimentary rocks) as well as basic igneous and metamorphic rocks create circumneutral and alkaline environments while siliceous sandstones, cherts and granites create acid ones. Additionally, shales are water-retaining while sandstones and limestones allow it to drain away. These factors combined with dynamic colluvial movements, rainwater leaching and capillary action interact powerfully with climate and topography, particularly in mountainous terrain, to induce habitat diversity on the most fundamental level. It is this complex of physical conditions, combined with biological factors, that govern the stability relations of the flora -- and in the end the fauna too. At times the relations are subtle and unexpected as I discovered while investigating the flora associated with a silica sandstone with a carbonate cement. In this case reaction with acid rainwater had formed a crust of secondary minerals which effectively sequestered the carbonate in the rock interior, allowing acidiphilic plants to grow on its surface. Thus an essentially thermodynamic process had moved the system along the pH axis to a microhabitat that may at first have seemed anomalous. In other cases another axis, that of transport and deposition, must be considered. The influence of this axis on the character of some tracts of old growth forest will be discussed in what follows.
It should be understood that much current research on forests is designed more to further their exploitation rather than ecological understanding or conservation. An example is the use of uniform study plots along line transects. This is of course an established method in many biological field investigations and has yielded much data of great ecological significance. As used in forestry however transects are frequently designed to avoid unusual topography and other features, pay little heed to non-timber species such as herbs and make few correlations between species and the mineral substrate. Data obtained are frequently averaged (a very un-ecological process!) and fed into statistics mills so microhabitats are obscured or ignored. Such data may be misleading because the uniformly-spaced plots or collection points may miss significant cryptic ecotones marked by species not recorded, not spatially located or which are averaged out of sight. By contrast, stability field analysis tells us that the precise location and relation to substrate and topography of each plant, whether woody or herbaceous, may be significant to the interpretation. Special and unusual topographic and substrate features are sought out to present the widest possible spectrum of habitats for comparison. In this way the roles of these features and associated species in the regional ecologic mosaic are revealed with the greatest economy. In many areas, including the Central Appalachians, these guidelines have generally not been followed and research has tended to concentrate on a few forest types of the many available, while detailed studies have tended to be confined to rare and unusual communities.ConclusionsAlthough the subject of old growth/primary forest has justifiably attracted attention of late, there appears to be considerable uncertainty and confusion regarding the origin of floral diversity in these forests. At a recent conference on old growth in eastern forests, emphasis was on developmental history and disturbance regimes. Although there were some attempts to evaluate the forest as a whole, trees were the focus of attention, while herbs and relation to the mineral substrate received little. Thus Williams (1997) called upon "site specific and regional historical and ecological factors" in an attempt to interpret herb layer species composition and richness in the old growth forest he studied, but did not specify what these factors were. It is contended here that attention to fundamental stability relations involving the mineral substrate and topography in such forests can contribute greatly to explaining variations in floral diversity and type.
In our inventory program Virginians for Wilderness have attempted to correlate the character of the herb layer, including bryophytes, with the mineral substrate and topographic factors in old growth/primary forest tracts in the Central Appalachians. Examples presented here include three tracts dominated by Eastern Hemlock (Tsuga candensis) and one of the mixed forests with Red Spruce (Picea rubens). The tracts with Hemlock include Dolly Ann Hollow in the George Washington National Forest and War Spur Hollow in the Mountain Lake Wilderness of the Jefferson National Forest, both in Virginia, and the Fanny Bennett Hemlock Grove of the Monongahela National Forest in West Virginia. The mixed forest with spruce is in the Gaudineer Scenic Area, also in the Monongahela.
All three of the Hemlock tracts also contain codominant Appalachian or Northern Hardwoods and the last two have Red Spruce as well, although the spruce in Fanny Bennett is largely in the understory. Trees in all three Hemlock tracts are large and vigorous but the character and richness of the shrub and herb layers vary considerably among them. Soils at Dolly Ann and War Spur are prevailingly acid and those of War Spur fall in the range of 2.9-4.4 (Adams and Stephensen, 1991). They are developed on Silurian Sandstone-dominated watersheds (Rader and Evans, 1993; Rader and Gathright, 1986). The situation at Fanny Bennett is more complex. While it is also underlain by sandstone, in this case Mississippian Pocono Formation, the same watershed up-slope from the Hemlock grove contains Greenbrier Limestone, also of Mississippian age (Cardwellet al, 1986). This limestone is in contact in varying degrees with two streams, the mainstream Sawmill Branch and a smaller branch of the latter. Both flow through Fanny Bennett.
Both Dolly Ann and War Spur have floras with acid soil affinities but the species differ somewhat because of elevational differences. Great Rhododendron (Rhododendron maximum), Mountain Laurel (Kalmia latifolia) and Blueberries (Vaccinium spp.) occur quite generally. Ground floras are sparse, lack diversity and are virtually confined to acid-demanding or tolerant plants such as Partridge Berry (Mitchella repens), Canada Mayflower (Maianthemum canadense) and Intermediate Shield Fern (Dryopteris intermedia). In contrast the character of shrub and herb floras of Fanny Bennett vary greatly in response to topographic position. Those under Hemlock on a southwest-facing slope are sparse and are comprised of acid-soil species including Painted Trillium (Trillium undulatum), Leucobryum cushion moss and liverwort Bazzania trilobata. These Hemlock stands grade upward into ericaceous oak-pine forest on a spur top. Soils on the the entire slope are derived from the Pocono sandstone.
The shrub and ground floras of the bottom-land streamside Hemlock forest are far more diverse and dense than those of either the adjacent slope forest or the streamside forests of Dolly Ann and War Spur. The difference is clearly a consequence of the access that the Fanny Bennett bottomland forests have to the Greenbrier limestone. Furthermore, even the degree of this access is reflected in the floras. In the bottomland along the small branch stream, which has the least access to the limestone, there is a rather rich flora with numerous acid-soil herbs such as Partridge Berry and Canada Mayflower as well as diverse bryophytes, including Plume Moss (Ptilium crista-castrensis), Mountain Fern Moss (Hylocomium splendens) and Bazzania. However these are joined by circumneutral-favoring species such as Wake Robin (Trillium erectum) and Wild Stonecrop (Sedum ternatum). In the mainstem bottom, herb diversity, density and luxuriance are substantially greater than along the branch stream and both woody and herbaceous plants are more indicative of neutral and even alkaline soils. These include, among many others, Jewelweed (Impatiens) of both species, Maidenhair Fern (Adiantum pedatum), Wood Nettle (Laportea canadensis), Slippery Elm (Ulmus rubra) and most indicative of all, the strongly calciphile Bulbiferous Bladder Fern (Cystopteris bulbifera).
In this example the relation between the flora and the mineral substrate is expanded to include the transport/deposition dimension, thus illustrating the dependence on the character of the enveloping topography as well. While in this case composition is to a large degree governed by alluvial processes, in other cases topographically-determined air drainage and accumulation may explain the presence of species from a different climatic zone. In the Central Appalachians this expressed as a mirroring effect, in which cold climate species such as Mountain Ash (Pyrus americana), Yellow Birch (Betula allegheniensis) and Canada Mayflower occur at both the highest elevations and in adjacent low valley flats but not on slopes in between.
The Gaudineer Scenic Area of old growth forest differs considerably from the three Hemlock-bearing tracts. Conifers, in this case Red Spruce, comprise less than 50% and dominance is shared by Yellow Birch, Red Maple (Acer rubrum), American Beech (Fagus grandifolia) with minor White Ash (Fraxinus americana) and Black Cherry (Prunus serotina). Topographically this forest occupies an upper slope with an easterly aspect and at 3900 ft (1200 meters) asl, is the highest of the four tracts discussed here. Bedrock, where observed by the writer, consists of very light gray conglomeratic sandstone of Carboniferous age, which decomposes to form a pebble-rich white sand. Because Spruce forms less than 50% of the tract, no continuous bryophyte/mor layer, such as characterizes pure spruce, has developed. Rather, in response to the rapid decay of Birch and Maple leaves, the organic soil layer is thin and discontinuous. According to Adams and Stephensen (1989) soil pH is in the range of 3.5. Although the red Spruce appears to show good reproduction in openings, the low rate of growth of both old and young spruce as measured by Adams and Stephensen supports their contention that this species appears to be in decline here, at least for now.
Although herb diversity at Gaudineer is considerably greater than at Dolly Ann, War Spur and the Fanny Bennett slope forest, this is probably a consequence of the dominantly deciduous canopy and the abundant springs around which many herbs are concentrated. The herb layer is dominated by acid-soil species such as White Wood Sorrel (Oxalis montana) and Intermediate Shield Fern. Where characteristic rich-soil species such as Wood Nettle do occur, they are stunted and unhealthy looking Thus the floral content and diversity of this tract of old growth also fits the pattern of substrate assertion and is appropriate to forest type and the climatic zone.
Stability field relations also bear directly on the frequently raised question regarding the existence of species particularly characteristic of, or which are confined to, old growth forest. Thus Martin (1992) suggested that Fraser's Sedge (Cymophyllus fraseri) might be an indicator of old growth conditions. However in every place I have observed this species it occupied second growth, albeit in rich soil. In one case the enveloping vegetation consisted of a young, acidic Spruce-Fir northern hardwood mixed forest while Fraser Sedge flourished in a narrow alluvial zone. Such occurrences illustrate the primacy of substrate and microhabitats established by whatever means over simple passage of time.
Nature's multidimensional space affords us a perspective which leads naturally to the stability field concept. Combined with classical ecological principles this concept offers a more comprehensive approach than has generally been applied to terrestrial ecosystems and forests in particular. Its principles elucidate such studies as those involving nutrient cycles, succession and the distribution of plants on both macro and micro scales because they are concerned with the most fundamental relation of plants to substrate. In her brilliant classic Deciduous Forests of Eastern North America, (1950), E. Lucy Braun sought to distinguish climatic, topographic and edaphic (soil) climaxes, believing that the first named would eventually prevail if enough time were available, and even today some forest ecologists still distinguish between the three. While there is little question that climate plays an important role in determining variations in forest type, particularly on a regional scale, bedrock and bedrock-derived materials have a dominant influence on smaller scales (Mueller & Mueller, 1997). Indeed, Braun's classic is rife with her discernment of this influence on a regional scale as well, particularly in her Western Mesophytic Forest Region. Examination of many forest types and stages of development in the Central Appalachians has led me to the conclusion that all stages of development from the earliest to climax result from the interplay of biologic, climatic and geologic factors and that all three factors are usually apparent in the vegetation and expressed in stability relations.AcknowledgementsStability field analysis of natural ecosystems and forests in particular also broadens the scientific basis of conservation biology through its incorporation of mineral and physical chemistry as well as analyses of climatic phenomena. In so doing we in a sense play catch-up to economic geology, which for years has been using plants as indicators of mineral deposits. We can do no less than exploit these and other tools available to protect our fast-dwindling ecosystems.
The writer is particularly indebted to Robert Hunsucker for many pleasurable days in the field and conversations and instruction regarding the Appalachian flora. He also appreciates the able word proceesing, advice and assistance of Gus Mueller and generous funding by Patagonia and Human-i-Tees.References
- Adams, Harold S. and Steven L. Stephensen (1989).
- "Old-growth Red Spruce Communities in the Mid-Appalachians," Vegetatio 85, 45-56
- Adams, Harold S. and Steven L. Stephenson (1991).
- High Elevation Coniferous Forests in Virginia," Virginia Journal of Science, 45 (4), 391-399
- Braun, E. Lucy (1950).
- Deciduous Forests of Eastern North America. Macmillan. New York.
- Cardwell, Dudley H., Robert B. Erwin, Herbert P. Woodward and Charles W. Lotz. compilers, (1968),
- Geologic Map of West Virginia, slightly revised 1986, West Virginia Geologic and Economic Survey
- Edsall, John T. and Jefferies Wyman (1958).
- Biophysical Chemistry, Vol. I, Academic Press, New York
- Hutchinson, G. E. ( 1965 ).
- The Ecological Theater and the Evolutionary Play, Yale Univ. Press, New Haven.
- Kretz, Ralph (1994).
- Metamorphic Crystallization, Wiley, New York
- Küchler, A. W. (1972).
- "Vegetation Indicators" in The Encyclopedia of Geochemistry and Environmental Sciences. Rhodes W. Fairbridge editor, Van Nostrand Reinhold Co., New York, 1238-1241.
- Martin, William H. (1992).
- "Characteristics of Old-Growth Mixed Mesophyte Forests," Natural Areas Journal, 12 (3), 127-135.
- Mueller, R.F. (1996).
- "Central Appalachian Plant Distributions and Forest Types or What A Walk in the Woods Can Tell You," Wild Earth 6 (1). 37-43.
- Mueller, R.F and Gus Mueller. (1997).
- "Geology in Reserve Design, an Example from the Folded Appalachians," Wild Earth 7 (2). 62-66.
- Rader, E.K. and T. M. Gathright II, editors (1986).
- Geologic Map of Giles County, Virginia, Virginia Division of Mineral Resources.
- Rader, E.K. and N. H. Evans, editors (1993).
- Geologic Map of Virginia - expanded explanation, Virginia Division of Mineral Resources.
- Williams, Charles E. (1997).
- "Herb Layer Species Composition and Richness in Eastern Old-Growth Forest: Local and Regional Variation," in Ancient Eastern US Old Growth Forests, Pattern, Process, Values and Management, Schedule and Abstracts for Science Agenda and Poster Session, June 5-8, 1997, Clarion, PA.
[1]For an excellent recent treatment on this subject, see the book by Kretz (1994).
When citing this paper please refer to Forests of the Central Appalachians Project, Virginians for Wilderness, 1998.